多倍体在植物中普遍存在,并且在几乎所有被子植物基因组的进化中发挥了重要作用。例如,许多驯化作物要么是多倍体(如小麦、咖啡、棉花和花生),要么是古多倍体(如水稻、玉米、大豆和高粱)。多倍化后,特别是在异源多倍体中,亚基因组可以通过再二倍化减少潜在的遗传不相容性,这是一个涉及快速基因丢失的过程。尽管二倍化在新基因和新物种的进化创新中发挥着重要作用,但驱动二倍化过程的进化力量及其潜在机制仍不清楚。
异源四倍体黍属禾本科稗科,是华北地区最早驯化的作物之一。然而,由于尚未确定祖先的二倍体祖细胞,在黍中亚基因组的鉴定和异源四倍体化时间估计具有挑战性,这限制了对这种古老的异源四倍体作物的起源和二倍体化的理解。
在最近的一项研究中,来自中国科学院(CAS)遗传与发育生物学研究所(IGDB)的陈明生教授团队重新组装了一个高质量的黍染色体基因组序列。
他们使用P.hallii作为中介,使用基于进化距离的方法将黍基因组分为两个亚基因组。亚基因组的成功鉴定为研究黍的异源多倍体进化提供了基础。此外,本研究中确定异源多倍体亚基因组的方法为其他未确认祖先二倍体祖细胞的异源多倍体研究提供了可靠的参考。
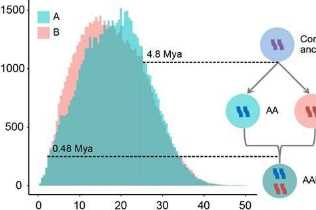
此外,他们计算了黍亚基因组之间的转座因子(TE)差异,以估计异源四倍体化事件的时间。他们的分析表明,这两个亚基因组在约480万年前出现分歧(Mya),而粟的异源四倍体化可能发生在约0.48Mya左右,这表明粟是相对较晚的异源四倍体。
比较分析表明,亚基因组B的大小大于亚基因组A,这是由于多倍化前亚基因组B的祖细胞中长末端重复逆转录转座子偏向积累所致。值得注意的是,富含转座子元素的亚基因组B中偏向突变的积累导致了更多的基因丢失。
此外,研究人员还透露,在两个亚基因组中共表达的同源基因和在一个亚基因组中表达的同源基因都没有显示出亚基因组表达优势的证据。然而,他们发现P.hallii中表达最少的基因在粟二倍体化过程中往往会丢失。
这项工作提供了一个二倍体化的例子,其中四倍体基因组经历了微妙的偏差分馏;然而,没有建立亚基因组表达优势。
这项名为“四倍体黍基因组二倍体化的偏向突变和基因丢失”的研究发表在植物杂志上。